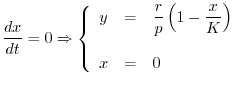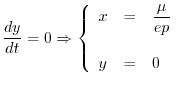Ci proponiamo ora di descrivere in maniera quantitativa l'interazione unidirezionale fra due comparti in cui una risorsa essenziale viene consumata da una popolazione. In particolare, vogliamo descrivere esplicitamente la dinamica accoppiata di prede e predatori, di piante ed erbivori, e di nutrienti e produttori primari.
Le ipotesi alla base dei modelli che illustreremo sono le seguenti:
Proposto indipendentemente nel 1924 dal demografo americano Alfred James
Lotka e nel 1926 dal matematico italiano Vito Volterra, quello di
Lotka-Volterra è stato storicamente il primo modello consumatore-risorsa. Nella
versione proposta dagli autori, in verità, l'accrescimento della risorsa
era maltusiano (
![]() ) e questa semplificazione matematica produceva
alcuni comportamenti singolari. Pur non essendo il modello molto realistico
neppure nella versione che presenteremo qui, perché
) e questa semplificazione matematica produceva
alcuni comportamenti singolari. Pur non essendo il modello molto realistico
neppure nella versione che presenteremo qui, perché ![]() non riflette la
realtà biologica, esso ha il grande pregio di permettere un'analisi semplice.
Rispetto alla notazione usata per la forma generale (15), le
funzioni
non riflette la
realtà biologica, esso ha il grande pregio di permettere un'analisi semplice.
Rispetto alla notazione usata per la forma generale (15), le
funzioni ![]() e
e ![]() del modello di Lotka-Volterra sono così
specificate:
del modello di Lotka-Volterra sono così
specificate:
Si otterrà dunque
Per studiare il comportamento del modello di Lotka-Volterra (16)
è bene iniziare dalla ricerca di eventuali soluzioni di equilibrio
della coppia di equazioni differenziali. Per questo basta imporre che si
annullino contemporaneamente le due derivate rispetto al tempo della
quantità ![]() di risorsa e della biomassa
di risorsa e della biomassa ![]() del consumatore. Le curve che
soddisfano la prima e la seconda delle due seguenti relazioni
del consumatore. Le curve che
soddisfano la prima e la seconda delle due seguenti relazioni
Se il predatore è sufficientemente efficace nell'interagire con la preda
(mortalità da fame contenuta e/o efficienza di conversione della
biomassa e coefficiente di predazione sufficientemente elevati in maniera
tale che
![]() ), le isocline (18) e (19) sono disposte come in Fig. 8.
Dal momento che in condizioni di equilibrio entrambe le derivate temporali
del modello (16) debbono annullarsi, ovvero debbono valere le condizioni
(17), gli equilibri del modello sono determinabili graficamente come le
intersezioni di una delle isocline della preda (tratteggiate in Fig. 8)
con una delle isocline del predatore (a tratto continuo in Fig. 8).
Come si evince dal grafico, gli equilibri sono in questo caso tre, e le loro
coordinate valgono:
), le isocline (18) e (19) sono disposte come in Fig. 8.
Dal momento che in condizioni di equilibrio entrambe le derivate temporali
del modello (16) debbono annullarsi, ovvero debbono valere le condizioni
(17), gli equilibri del modello sono determinabili graficamente come le
intersezioni di una delle isocline della preda (tratteggiate in Fig. 8)
con una delle isocline del predatore (a tratto continuo in Fig. 8).
Come si evince dal grafico, gli equilibri sono in questo caso tre, e le loro
coordinate valgono:
![$ \bar{X}_0=
\left[\begin{array}{c}
0\\
\\
0
\end{array}\right]
$](img517.gif) |
![$ \bar{X}_1=
\left[\begin{array}{c}
K\\
\\
0
\end{array}\right]
$](img518.gif) |
![$ \bar{X}_2=
\left[\begin{array}{c}
\dfrac{\mu}{ep}\\
\\
\dfrac{r}{p}\left(1-\frac{x}{K}\right)
\end{array}\right]
$](img519.gif) |
![\includegraphics[scale=1]{lotka-volterra}](img520.gif) |
L'equilibrio ![]() , che
solitamente viene detto banale, corrisponde all'estinzione ovvero
all'assenza simultanea di prede e predatori. L'equilibrio
, che
solitamente viene detto banale, corrisponde all'estinzione ovvero
all'assenza simultanea di prede e predatori. L'equilibrio ![]() corrisponde all'assenza di consumatori, cosicché la
risorsa ha come equilibrio la sua capacità
portante. La terza soluzione di regime è quella più interessante,
perché corrisponde alla coesistenza contemporanea di prede e predatori.
Si noti però che, contrariamente ai primi due equilibri, tale soluzione
non esiste sempre: il valore di
corrisponde all'assenza di consumatori, cosicché la
risorsa ha come equilibrio la sua capacità
portante. La terza soluzione di regime è quella più interessante,
perché corrisponde alla coesistenza contemporanea di prede e predatori.
Si noti però che, contrariamente ai primi due equilibri, tale soluzione
non esiste sempre: il valore di ![]() in
in ![]() può assumere
valore negativo se
può assumere
valore negativo se
![]() , cioè se, come dicevamo prima,
il predatore è particolarmente inefficiente.
, cioè se, come dicevamo prima,
il predatore è particolarmente inefficiente.
Non tutti e tre gli equilibri trovati sono tuttavia stabili. Formalmente, un
metodo rigoroso e semplice per decidere della stabilità di un equilibrio
in un modello non lineare consiste nel determinare il segno della parte
reale degli autovalori del sistema linearizzato in un intorno del punto di
equilibrio. Per chi non avesse dimestichezza con tali metodi quantitativi,
diciamo che il risultato dell'analisi di stabilità porta a concludere
che gli equilibri ![]() e
e ![]() sono instabili,
mentre
sono instabili,
mentre ![]() è stabile.
è stabile.
Un'analisi qualitativa dell'andamento delle traiettorie del modello, che si
basa sulle isocline prima determinate, può essere effettuata come segue.
Ciascuna delle isocline non banali, ovvero la prima delle (18) e la prima
delle (19), divide il primo quadrante dello spazio di stato ![]() in due
parti. Ricordiamo che le isocline rappresentano il luogo dei punti nello
spazio di stato in cui il vettore tangente alla traiettoria ha componente
orizzontale o verticale nulla. In particolare, l'isoclina non banale
tratteggiata, la prima delle (18), viene attraversata verticalmente
poiché soddisfa la condizione
in due
parti. Ricordiamo che le isocline rappresentano il luogo dei punti nello
spazio di stato in cui il vettore tangente alla traiettoria ha componente
orizzontale o verticale nulla. In particolare, l'isoclina non banale
tratteggiata, la prima delle (18), viene attraversata verticalmente
poiché soddisfa la condizione ![]() , mentre quella a tratto continuo, la
prima delle (19), viene attraversata orizzontalmente. Si noti che la
componente orizzontale del vettore tangente alla traiettoria per tutti i
punti dello spazio di stato che stanno sopra l'isoclina non banale
tratteggiata ha verso orientato a sinistra. Ciò si vede dal fatto che,
al limite di
, mentre quella a tratto continuo, la
prima delle (19), viene attraversata orizzontalmente. Si noti che la
componente orizzontale del vettore tangente alla traiettoria per tutti i
punti dello spazio di stato che stanno sopra l'isoclina non banale
tratteggiata ha verso orientato a sinistra. Ciò si vede dal fatto che,
al limite di ![]() molto grandi, la prima delle (16) ha segno negativo. Per
esclusione, ne deriva che tutti i punti dello spazio di stato che stanno
sotto l'isoclina non banale tratteggiata hanno componente orizzontale del
vettore tangente diretta verso destra. Analogo ragionamento si può
svolgere per analizzare il verso della componente verticale del vettore
tangente alla traiettoria. Per questo, è sufficiente valutare il segno
della seconda delle (16) per uno dei punti a destra dell'isoclina non
banale a tratto continuo. Dalla seconda delle (16) si evince che, al
limite di
molto grandi, la prima delle (16) ha segno negativo. Per
esclusione, ne deriva che tutti i punti dello spazio di stato che stanno
sotto l'isoclina non banale tratteggiata hanno componente orizzontale del
vettore tangente diretta verso destra. Analogo ragionamento si può
svolgere per analizzare il verso della componente verticale del vettore
tangente alla traiettoria. Per questo, è sufficiente valutare il segno
della seconda delle (16) per uno dei punti a destra dell'isoclina non
banale a tratto continuo. Dalla seconda delle (16) si evince che, al
limite di ![]() molto grandi, il vettore tangente alla traiettoria deve avere
componente verticale diretta verso l'alto. Conseguentemente, i punti a
sinistra dell'isoclina verticale hanno componente verticale del vettore
tangente alla traiettoria diretto verso il basso. Dal momento che le
isocline non banali sono in questo caso rette che si intersecano in un punto
(l'equilibrio stabile
molto grandi, il vettore tangente alla traiettoria deve avere
componente verticale diretta verso l'alto. Conseguentemente, i punti a
sinistra dell'isoclina verticale hanno componente verticale del vettore
tangente alla traiettoria diretto verso il basso. Dal momento che le
isocline non banali sono in questo caso rette che si intersecano in un punto
(l'equilibrio stabile ![]() ), il primo quadrante risulta suddiviso in quattro regioni:
(a), (b), (c) e (d). Grazie all'analisi
qualitativa appena effettuata, siamo in grado di determinare quantomeno
direzione e verso del vettore tangente alla traiettoria in ognuno dei punti
di tali regioni. L'andamento di una traiettoria coerente con l'analisi
effettuata e che si origina nel punto
), il primo quadrante risulta suddiviso in quattro regioni:
(a), (b), (c) e (d). Grazie all'analisi
qualitativa appena effettuata, siamo in grado di determinare quantomeno
direzione e verso del vettore tangente alla traiettoria in ognuno dei punti
di tali regioni. L'andamento di una traiettoria coerente con l'analisi
effettuata e che si origina nel punto ![]() è mostrato in Fig.8. Esso fa
capire in maniera intuitiva che effettivamente l'equilibrio
è mostrato in Fig.8. Esso fa
capire in maniera intuitiva che effettivamente l'equilibrio ![]() ``attrae'' verso di
sè le traiettorie, cioè è stabile.
``attrae'' verso di
sè le traiettorie, cioè è stabile.
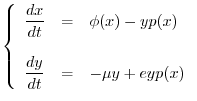
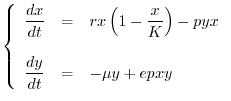
![$\displaystyle \left\{\begin{array}{lll}
\dfrac{dx}{dt} & = & \left[r\left(1-\d...
...&\\
\dfrac{dy}{dt} & = & \left[-\mu + epx\right]y = 0 \\
\end{array}\right.$](img514.gif)