Comunità acquatiche
La profondità alla quale la luce del sole riesce a penetrare nell'acqua di un lago o di un
mare è un fattore critico per definire la zona entro cui avviene la produzione primaria,
che nelle comunità acquatiche è essenzialmente a carico del fitoplankton (salvo che in
corpi d'acqua poco profondi, dove la vegetazione radicata può essere predominante). L'acqua
assorbe rapidamente la radiazione solare, per cui la quantità di radiazione che arriva alle
diverse quote diminuisce con la profondità. Questa diminuzione è descritta
ragionevolmente bene dalla legge di Beer. Indichiamo con ![]() la profondità
misurata a partire dal pelo libero dell'acqua e con
la profondità
misurata a partire dal pelo libero dell'acqua e con ![]() la quantità di radiazione solare
per unità di area che arriva alla profondità
la quantità di radiazione solare
per unità di area che arriva alla profondità ![]() . La percentuale di radiazione solare
che viene assorbita da uno strato d'acqua di altezza dz è approssimativamente
proporzionale all'altezza dello strato attraverso una costante
. La percentuale di radiazione solare
che viene assorbita da uno strato d'acqua di altezza dz è approssimativamente
proporzionale all'altezza dello strato attraverso una costante ![]() , detta coefficiente di
estinzione.
, detta coefficiente di
estinzione. ![]() ovviamente dipende dalla trasparenza dell'acqua ed è tanto più alto
quanto più l'acqua è torbida. Possiamo scrivere il seguente bilancio per uno
straterello d'acqua di altezza dz, di area unitaria e situato alla profondità
ovviamente dipende dalla trasparenza dell'acqua ed è tanto più alto
quanto più l'acqua è torbida. Possiamo scrivere il seguente bilancio per uno
straterello d'acqua di altezza dz, di area unitaria e situato alla profondità ![]() :
:
![]() +dz) =
+dz) = ![]() - k dz I(
- k dz I(![]() .
.
Passando al limite otteniamo la legge di Beer
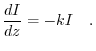
Tuttavia la PPL alle diverse profondità non segue in generale una legge di decremento
esponenziale: il massimo della produzione primaria può verificarsi diversi metri al di
sotto della superficie dell'acqua. Questo avviene perchè troppa luce inibisce il processo
della fotosintesi: in generale l'efficienza fotosintetica non è una costante indipendente
da ![]() , ma è una funzione decrescente di
, ma è una funzione decrescente di ![]() . A seconda dei valori del coefficiente di
estinzione, della luce incidente sulla superficie dell'acqua e dell'andamento dell'efficienza
fotosintetica si possono avere situazioni molto diversificate nei corpi d'acqua. La figura
2 riporta le produzioni primarie lorde in funzione della profondità per
tre diversi laghi californiani: Clear Lake (a dispetto del nome!) è un lago
eutrofico, cioè con elevata produzione primaria, scarsa penetrazione della luce,
fotosintesi che si esaurisce nei primi 4 metri, Lake Tahoe all'opposto è un lago alpino
oligotrofico, cioè con bassa produzione primaria, luce che penetra fino a 60 m,
massimo della fotosintesi intorno ai 15 m, mentre Castle Lake ha PPL intermedia, un massimo ben
marcato della fotosintesi intorno agli 8 m e radiazione solare che si esaurisce entro i 30 m.
. A seconda dei valori del coefficiente di
estinzione, della luce incidente sulla superficie dell'acqua e dell'andamento dell'efficienza
fotosintetica si possono avere situazioni molto diversificate nei corpi d'acqua. La figura
2 riporta le produzioni primarie lorde in funzione della profondità per
tre diversi laghi californiani: Clear Lake (a dispetto del nome!) è un lago
eutrofico, cioè con elevata produzione primaria, scarsa penetrazione della luce,
fotosintesi che si esaurisce nei primi 4 metri, Lake Tahoe all'opposto è un lago alpino
oligotrofico, cioè con bassa produzione primaria, luce che penetra fino a 60 m,
massimo della fotosintesi intorno ai 15 m, mentre Castle Lake ha PPL intermedia, un massimo ben
marcato della fotosintesi intorno agli 8 m e radiazione solare che si esaurisce entro i 30 m.
![\includegraphics[width=0.8\linewidth]{lakes}](img500.gif) |
Gli esempi di figura 2 mostrano come il fattore luce possa spiegare
l'andamento della produzione primaria nel medesimo lago ma non possa spiegare come mai i tre
laghi sono differenti tra di loro avendo PPL medie che differiscono di ordini di grandezza. La
ragione fondamentale è la diversa disponibilità di sali nutrienti, cioè di
sali contenenti tutti quegli elementi che permettono alle alghe di costruire la propria
biomassa. In ambiente acquatico, le risorse limitanti - ovvero disponibili in minore quantità rispetto alle necessità del fitoplankton - sono in genere azoto e fosforo. In
particolare sono carenti nitrati e fosfati, che sono i composti di ![]() e
e ![]() effettivamente
assorbibili dalle alghe. Ciò è particolarmente vero per le acque superficiali (si veda
la Fig. 3): le acque profonde, che ricevono buona parte del materiale organico
morto che lentamente decade e in cui non c'è assorbimento da parte delle alghe, sono invece
ricche di nutrienti. D'altra parte la fotosintesi non può che avvenire in superficie e
quindi la scarsità dei nutrienti rende gli oceani dei deserti a tutti gli effetti.
effettivamente
assorbibili dalle alghe. Ciò è particolarmente vero per le acque superficiali (si veda
la Fig. 3): le acque profonde, che ricevono buona parte del materiale organico
morto che lentamente decade e in cui non c'è assorbimento da parte delle alghe, sono invece
ricche di nutrienti. D'altra parte la fotosintesi non può che avvenire in superficie e
quindi la scarsità dei nutrienti rende gli oceani dei deserti a tutti gli effetti.
![\includegraphics[scale=0.8]{nitrati}](img501.gif) |
Esistono tuttavia delle eccezioni nello spazio e nel tempo a questa situazione. In particolare in oceano aperto esistono zone di upwelling, ovvero di risalita delle acque profonde ricche di nutrienti, dovute al regime generale delle correnti marine. Queste zone oceaniche (ad es. al largo delle coste del Peru e lungo una buona parte dell'Antartide) sono particolarmente produttive. Tutta la catena trofica ne riceve un impulso; le popolazioni di pesci e crostacei sono particolarmente abbondanti e sostengono un'importante industria della pesca. Nei laghi profondi delle regioni temperate invece si verifica una volta o due volte all'anno il fenomeno di rottura della stratificazione delle acque con conseguente ricircolo dei nutrienti presenti nelle acque profonde (si veda la Fig. 4).
In che cosa consiste il fenomeno? Poiché l'acqua ha la sua massima densità a 4![]() C,
in inverno la superficie dei laghi è ghiacciata o comunque a temperature prossime allo
zero e l'acqua più fredda galleggia su quella meno fredda (stratificazione invernale).
Quando sopraggiunge la primavera, l'acqua superficiale si riscalda, diventa più densa
e precipita negli strati sottostanti, che tendono a risalire e a scaldarsi. Con l'aiuto
dei venti questi movimenti, che di per sé creerebbero una situazione omogenea di distribuzione
termica, vengono accentuati e l'acqua di fondo tende a risalire anch'essa portando con sè nutrienti.
Col procedere della primavera e l'arrivo dell'estate, tutta l'acqua diventa più calda
di 4
C,
in inverno la superficie dei laghi è ghiacciata o comunque a temperature prossime allo
zero e l'acqua più fredda galleggia su quella meno fredda (stratificazione invernale).
Quando sopraggiunge la primavera, l'acqua superficiale si riscalda, diventa più densa
e precipita negli strati sottostanti, che tendono a risalire e a scaldarsi. Con l'aiuto
dei venti questi movimenti, che di per sé creerebbero una situazione omogenea di distribuzione
termica, vengono accentuati e l'acqua di fondo tende a risalire anch'essa portando con sè nutrienti.
Col procedere della primavera e l'arrivo dell'estate, tutta l'acqua diventa più calda
di 4![]() C e quindi l'acqua più calda galleggia su quella meno calda. Inoltre, poiché, come
abbiamo visto, la radiazione solare viene praticamente tutta assorbita dallo strato superficiale, a partire da
una certa profondità la temperatura non varia quasi più mentre lo strato superficiale continua
a riscaldarsi. Lo strato superficiale, ben illuminato, caratterizzato da acque calde, si chiama epilimnio,
mentre le acque più fredde sottostanti prendono il nome di ipolimnio.
C e quindi l'acqua più calda galleggia su quella meno calda. Inoltre, poiché, come
abbiamo visto, la radiazione solare viene praticamente tutta assorbita dallo strato superficiale, a partire da
una certa profondità la temperatura non varia quasi più mentre lo strato superficiale continua
a riscaldarsi. Lo strato superficiale, ben illuminato, caratterizzato da acque calde, si chiama epilimnio,
mentre le acque più fredde sottostanti prendono il nome di ipolimnio.
![\includegraphics[scale=0.20]{stratificazione}](img502.gif) |
La zona di rapida variazione della temperatura, che divide i due strati in estate, si chiama
termoclinio. La figura 5 mostra il formarsi della netta
stratificazione termica estiva nel Lago Harsha in Ohio (USA). Con l'arrivo dell'autunno le acque superficiali diventano fredde più
rapidamente e, quando raggiungono una temperatura inferiore a quella dell'ipolimnio, tendono a
precipitare sul fondo distruggendo il termoclinio e la temperatura tende ad uniformarsi al
valore di 4![]() C. In inverno si riforma la stratificazione opposta a quella dell'estate con
acque più fredde che galleggiano al di sopra di quelle meno fredde. I due ricircoli spiegano
perché le fioriture algali, cioè periodi di elevata produzione primaria, si verificano
prevalentemente in primavera e in autunno. Il ricircolo autunnale di acque è di solito più
importante di quello primaverile perché la stratificazione estiva è molto più marcata di
quella invernale. Tuttavia esso avviene quando le temperature sono già piuttosto basse e la
fotosintesi non è quindi molto favorita. Per questa ragione la fioritura algale autunnale non
si verifica sempre.
C. In inverno si riforma la stratificazione opposta a quella dell'estate con
acque più fredde che galleggiano al di sopra di quelle meno fredde. I due ricircoli spiegano
perché le fioriture algali, cioè periodi di elevata produzione primaria, si verificano
prevalentemente in primavera e in autunno. Il ricircolo autunnale di acque è di solito più
importante di quello primaverile perché la stratificazione estiva è molto più marcata di
quella invernale. Tuttavia esso avviene quando le temperature sono già piuttosto basse e la
fotosintesi non è quindi molto favorita. Per questa ragione la fioritura algale autunnale non
si verifica sempre.
![\includegraphics[width=.75\linewidth]{LakeHarsha}](img503.gif) |
Tra azoto e fosforo, è sostanzialmente il fosforo ad essere il fattore
limitante la produzione primaria in ambienti acquatici (si veda la Fig. 6, che risulta da una
statistica effettuata su diversi laghi). Infatti, in condizioni non
perturbate dall'uomo, l'apporto di fosforo ai corpi d'acqua è piuttosto
piccolo, poiché vi è poco fosforo non solo nelle precipitazioni, ma
anche negli afflussi superficiali, essendo il fosforo trattenuto dalla
vegetazione e da reazioni chimiche nel suolo. È raro che l'azoto diventi
limitante: una diminuzione della concentrazione di nitrati può
verificarsi a causa del processo di denitrificazione, cioè di riduzione
dello ione nitrato ad azoto atmosferico operato da alcuni batteri
specializzati, ma in questo caso quello che succede è che nella
comunità fitoplantonica si sviluppano particolari alghe, i cianobatteri
o alghe azzurro-verdi, che sono in grado di fissare direttamente l'azoto.
Smith (1983) trovò che i cianobatteri sono frequenti nella comunità
fitoplantonica quando il rapporto ![]() /
/![]() è inferiore a 29. Quando gli
scarichi inquinanti nei corpi d'acqua apportano notevoli quantità di
fosforo, l'azoto dovrebbe diventare il fattore limitante, ma in realtà
quello che succede è che la PPN aumenta, perché si sviluppano le
alghe azzurro-verdi.
è inferiore a 29. Quando gli
scarichi inquinanti nei corpi d'acqua apportano notevoli quantità di
fosforo, l'azoto dovrebbe diventare il fattore limitante, ma in realtà
quello che succede è che la PPN aumenta, perché si sviluppano le
alghe azzurro-verdi.
![\includegraphics[width=0.5\linewidth]{fosfPPN}](img504.gif) |
Comunità terrestri
A parità di condizioni climatiche la disponibilità di nutrienti
è il fattore che determina la produzione primaria anche in ambiente
terrestre. Molto frequentemente è l'azoto il nutriente limitante:
l'aggiunta di fertilizzanti azotati nel suolo non solo stimola la crescita
delle piante nei sistemi agricoli, ma quasi sempre aumenta la
produttività anche delle foreste. Naturalmente negli ecosistemi naturali
l'azoto non è artificialmente aggiunto dall'uomo. Esso è tuttavia
quasi sempre di origine biologica. Infatti, nonostante l'enorme quantità
di azoto presente nell'atmosfera, è solo sotto forma di ammoniaca
(NH![]() o di nitrati (NO
o di nitrati (NO![]() che esso può venire
utilizzato dalla grande maggioranza delle piante.
che esso può venire
utilizzato dalla grande maggioranza delle piante.
Tra le piante terrestri solo i legumi hanno la capacità di fissare l'azoto atmosferico. In realtà questa capacità non è intrinseca, ma dovuta al fatto che nei noduli delle radici dei legumi vivono in simbiosi batteri del genere Rhizobium, che utilizzando l'energia della pianta sono in grado di trasformare azoto atmosferico in ammoniaca. In Fig. 7 sono mostrati a titolo esemplificativo i noduli di Rhizobium naturalmente contenuti nel sistema radicale di una pianta di piselli. La pratica di piantare legumi per ripristinare la fertilità di suoli eccessivamente impoveriti è nota agli agricoltori da tempo immemorabile. Oltre a Rhizobium, numerosi altri batteri viventi in diversi tipi di suoli sono in grado di fissare l'azoto atmosferico e di trasformarlo o in ammoniaca o in nitrati (processo di nitrificazione), rendendolo così disponibile per la produzione primaria.
![\includegraphics[width=0.2\linewidth]{rhizobium}](img506.gif)