Abbiamo visto che il bilancio energetico di una catena alimentare può
facilmente essere descritto attraverso un sistema dinamico in cui il
contenuto energetico immagazzinato in ogni singolo livello trofico è
rappresentato dalla biomassa totale del comparto corrispondente. Per
esempio, in una catena di pascolo formata da produttori primari (![]() ,
produttori secondari (
,
produttori secondari (![]() = erbivori), consumatori di primo e secondo livello
(
= erbivori), consumatori di primo e secondo livello
(![]() e
e ![]() = carnivori) si può scrivere il modello
(14) di pag.
= carnivori) si può scrivere il modello
(14) di pag. ![[*]](crossref.gif) che, per praticità,
riscriviamo di seguito
che, per praticità,
riscriviamo di seguito
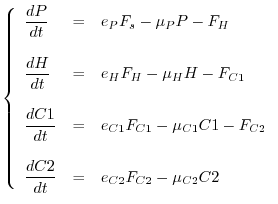
Ci limiteremo dunque a descrivere semplicemente la dinamica della più semplice catena trofica, costituita da una risorsa (o una popolazione di prede) e da una popolazione di consumatori (predatori). Nel caso che il consumatore sia un carnivoro i modelli corrispondenti vengono chiamati modelli preda-predatore, mentre nel caso che il consumatore sia un erbivoro si parla di modelli di pascolamento. È utile introdurre modelli consumatore-risorsa anche nel caso dei soli produttori primari, il cui sostentamento è naturalmente legato alla disponibilità di sali nutrienti, che costituiscono un comparto abiotico.
[r]![\includegraphics[scale=0.1]{liebig}](img489.gif) Come abbiamo già detto nel Capitolo 4, nonostante l'energia
immagazzinata dalle piante attraverso il processo di fotosintesi sia fornita
dal sole, non è in generale il fattore luce a determinare la produzione
primaria netta (che costituisce il flusso di energia immagazzinata nel
comparto delle piante) bensì la diversa disponibilità di sali
nutrienti, cioè di sali contenenti tutti gli elementi che permettono
alle piante di costruire la propria biomassa. Nel caso dei produttori
primari, la risorsa non è un'altra popolazione ad un inferiore livello
trofico, ma un composto inorganico. A tale proposito, è bene chiarire
che le piante, come tutti gli esseri viventi, richiedono diversi elementi
per il proprio sostentamento (quali azoto, fosforo, carbonio, manganese, ferro,
ecc.): per costruire la propria biomassa, esse debbono quindi assorbire un
gran numero di sali nutrienti. Tuttavia, come mise in evidenza
nell'Ottocento il chimico e agronomo Justus von Liebig (1808-1873, qui sopra),
è in generale uno solo il sale nutriente che limita la crescita
di una pianta e precisamente quello che è presente in minore
quantità nel terreno o nell'acqua rispetto alle necessità della
pianta (legge del minimo di Liebig). Per esempio: se la biomassa secca di
una pianta è costituita per l'1.5% da azoto e per lo 0.5% da
fosforo e il terreno contiene 200 unità di azoto e 100 unità di
fosforo è l'azoto che limita la crescita della pianta, poiché
Come abbiamo già detto nel Capitolo 4, nonostante l'energia
immagazzinata dalle piante attraverso il processo di fotosintesi sia fornita
dal sole, non è in generale il fattore luce a determinare la produzione
primaria netta (che costituisce il flusso di energia immagazzinata nel
comparto delle piante) bensì la diversa disponibilità di sali
nutrienti, cioè di sali contenenti tutti gli elementi che permettono
alle piante di costruire la propria biomassa. Nel caso dei produttori
primari, la risorsa non è un'altra popolazione ad un inferiore livello
trofico, ma un composto inorganico. A tale proposito, è bene chiarire
che le piante, come tutti gli esseri viventi, richiedono diversi elementi
per il proprio sostentamento (quali azoto, fosforo, carbonio, manganese, ferro,
ecc.): per costruire la propria biomassa, esse debbono quindi assorbire un
gran numero di sali nutrienti. Tuttavia, come mise in evidenza
nell'Ottocento il chimico e agronomo Justus von Liebig (1808-1873, qui sopra),
è in generale uno solo il sale nutriente che limita la crescita
di una pianta e precisamente quello che è presente in minore
quantità nel terreno o nell'acqua rispetto alle necessità della
pianta (legge del minimo di Liebig). Per esempio: se la biomassa secca di
una pianta è costituita per l'1.5% da azoto e per lo 0.5% da
fosforo e il terreno contiene 200 unità di azoto e 100 unità di
fosforo è l'azoto che limita la crescita della pianta, poiché
![\includegraphics[scale=0.7]{comparti}](img488.gif)