Il processo di fotosintesi clorofilliana è fondamentale per l'esistenza
della materia vivente sulla terra. Esso permette di trasferire il carbonio
dalla sua forma ossidata, l'anidride carbonica CO![]() , presente
nell'atmosfera, alla sua forma ridotta (carboidrati), presente nei tessuti
delle piante, le quali costituiscono in peso più del 99% della
materia vivente sulla nostra terra. A questo proposito si tenga presente che
le molecole biologiche, essenzialmente costituite da C, H, N, O, P e S, sono
caratterizzate da legami relativamente deboli, o, come si dice, ridotti,
cioè ricchi di elettroni. Secondo le leggi della termodinamica le
reazioni chimiche procedono spontaneamente verso situazioni di minore
energia libera, caratterizzate da forti legami chimici e da massimo
disordine. La materia vivente è in un certo senso una contraddizione a
queste leggi in quanto le molecole degli organismi viventi continuano ad
esistere in presenza di un forte agente ossidante come l'ossigeno
atmosferico che tende ad acquisire elettroni. L'evoluzione verso una
situazione di equilibrio termodinamico comporterebbe la formazione di
CO
, presente
nell'atmosfera, alla sua forma ridotta (carboidrati), presente nei tessuti
delle piante, le quali costituiscono in peso più del 99% della
materia vivente sulla nostra terra. A questo proposito si tenga presente che
le molecole biologiche, essenzialmente costituite da C, H, N, O, P e S, sono
caratterizzate da legami relativamente deboli, o, come si dice, ridotti,
cioè ricchi di elettroni. Secondo le leggi della termodinamica le
reazioni chimiche procedono spontaneamente verso situazioni di minore
energia libera, caratterizzate da forti legami chimici e da massimo
disordine. La materia vivente è in un certo senso una contraddizione a
queste leggi in quanto le molecole degli organismi viventi continuano ad
esistere in presenza di un forte agente ossidante come l'ossigeno
atmosferico che tende ad acquisire elettroni. L'evoluzione verso una
situazione di equilibrio termodinamico comporterebbe la formazione di
CO![]() , H
, H![]() O e NO
O e NO
![]() . In effetti questo è proprio
quello che succede quando gli organismi muoiono. Le piante, per mantenersi
vive, devono continuamente processare l'energia che viene dal sole per
rimanere in una situazione di non equilibrio termodinamico caratterizzato da
strutture altamente ordinate tenute insieme da deboli legami chimici.
. In effetti questo è proprio
quello che succede quando gli organismi muoiono. Le piante, per mantenersi
vive, devono continuamente processare l'energia che viene dal sole per
rimanere in una situazione di non equilibrio termodinamico caratterizzato da
strutture altamente ordinate tenute insieme da deboli legami chimici.
![\includegraphics[scale=1]{foglia}](img156.gif) |
La fotosintesi avviene nei cloroplasti delle cellule delle foglie delle piante verdi. La struttura della sezione di una foglia è mostrata in Fig. 1. In essa si riconoscono le seguenti componenti:
La fotosintesi è resa possibile da un pigmento, la clorofilla, che assorbe l'energia solare, specialmente le lunghezze d'onda rosse e blu, e la trasferisce a diverse reazioni chimiche, che comportano l'utilizzo di alcuni specifici enzimi. Questi enzimi permettono da una parte di spezzare le molecole d'acqua e di rilasciare ossigeno nell'atmosfera, dall'altra di utilizzare idrogeno e anidride carbonica per costruire molecole di carboidrati. In forma semplificata possiamo riassumere la fotosintesi nella maniera seguente
Per mantenere le complesse molecole biologiche che ne costituiscono i tessuti le piante, come tutti gli esseri viventi, devono spendere parte dell'energia che hanno accumulato. Questi processi vengono chiamati metabolici e comportano la perdita di energia sotto forma di calore (vedi la Fig. 2). La respirazione è proprio l'opposto della fotosintesi e, in forma semplificata, possiamo scrivere che è data da
I processi di fotosintesi e respirazione sono sintetizzati schematicamente in
Fig. 2. Essa mostra due importanti cicli di feedback, quello della CO![]() e
quello dell'acqua. La parte sinistra del disegno mostra l'effetto indotto
dalla luce. Promuovendo la fotosintesi, la luce causa una diminuzione del livello di CO
e
quello dell'acqua. La parte sinistra del disegno mostra l'effetto indotto
dalla luce. Promuovendo la fotosintesi, la luce causa una diminuzione del livello di CO![]() nella foglia. La risposta della foglia consiste nell'aumentare la
concentrazione di ioni potassio (K
nella foglia. La risposta della foglia consiste nell'aumentare la
concentrazione di ioni potassio (K![]() nelle cellule di guardia;
l'acqua, trascinata per osmosi, gonfia le cellule di guardia facendo
così aprire gli stomi. Nella parte di destra di Fig. 2,
invece, è rappresentato l'effetto dello stress idrico.
Quando dalla superficie fogliare evapora più acqua di quella che
può arrivare dalle radici, la foglia causa una diminuzione del livello
di ioni potassio nelle cellule di guardia, facendo così uscire acqua
(sempre per osmosi) e provocando la chiusura degli stomi. Se il tasso di
traspirazione è particolarmente elevato, l'acqua evapora direttamente
dalle cellule di guardia provocando comunque la chiusura degli stomi.
nelle cellule di guardia;
l'acqua, trascinata per osmosi, gonfia le cellule di guardia facendo
così aprire gli stomi. Nella parte di destra di Fig. 2,
invece, è rappresentato l'effetto dello stress idrico.
Quando dalla superficie fogliare evapora più acqua di quella che
può arrivare dalle radici, la foglia causa una diminuzione del livello
di ioni potassio nelle cellule di guardia, facendo così uscire acqua
(sempre per osmosi) e provocando la chiusura degli stomi. Se il tasso di
traspirazione è particolarmente elevato, l'acqua evapora direttamente
dalle cellule di guardia provocando comunque la chiusura degli stomi.
![\includegraphics[scale=1]{guardia}](img161.gif) |
Naturalmente l'accumulo di materia vivente nelle piante (cioè la produzione primaria) è possibile perché la fotosintesi e la respirazione non si compensano, ovvero l'energia catturata attraverso il processo di fotosintesi è maggiore dell'energia persa col processo di respirazione. E questo nonostante che di notte avvenga solo la respirazione. La differenza di energia viene immagazzinata come tessuto vegetale che, tra l'altro, costituisce il cibo per gli erbivori (secondo livello della catena alimentare) e rende quindi possibile il trasferimento di energia agli organismi eterotrofi. Più precisamente, con riferimento ad un determinato lasso di tempo, si definisce
| Produzione primaria lorda (PPL) | = | energia acquisita con la fotosintesi |
| Produzione primaria netta (PPN) | = | PPL - energia persa con la respirazione |
La misura delle produzioni primarie, sia lorda sia netta, costituisce un
argomento di grande importanza, in quanto permette di capire su quale flusso
di energia possa contare un determinato ecosistema ai fini del proprio
funzionamento. I metodi sono parecchi e differiscono a seconda
dell'ecosistema considerato. Per le piante terrestri una maniera diretta
è quella di misurare i cambiamenti di concentrazione di CO![]() nell'aria attorno alle piante. A questo fine si racchiude una pianta o parte
di essa in un involucro trasparente e si misurano i flussi di CO
nell'aria attorno alle piante. A questo fine si racchiude una pianta o parte
di essa in un involucro trasparente e si misurano i flussi di CO![]() .
Infatti esiste un ben preciso rapporto tra la quantità di energia solare
catturata da una pianta e la quantità di anidride carbonica scambiata
con l'atmosfera. Più precisamente, se consideriamo la reazione
semplificata della fotosintesi come espressa in moli, allora l'energia
solare necessaria perché avvenga la reazione è esattamente 709 kcal.
Perciò lo scambio di 6 moli di CO
.
Infatti esiste un ben preciso rapporto tra la quantità di energia solare
catturata da una pianta e la quantità di anidride carbonica scambiata
con l'atmosfera. Più precisamente, se consideriamo la reazione
semplificata della fotosintesi come espressa in moli, allora l'energia
solare necessaria perché avvenga la reazione è esattamente 709 kcal.
Perciò lo scambio di 6 moli di CO![]() (circa 134 litri a pressione e
temperatura standard) indica che sono state assorbite 709 kcal. Ricordando
che il peso atomico del carbonio è 12 possiamo anche dire che lo scambio
di 6
(circa 134 litri a pressione e
temperatura standard) indica che sono state assorbite 709 kcal. Ricordando
che il peso atomico del carbonio è 12 possiamo anche dire che lo scambio
di 6![]() 12 = 72 gC equivale ad introitare 709 kcal. Naturalmente la
stessa equivalenza si ha anche nel processo di respirazione.
12 = 72 gC equivale ad introitare 709 kcal. Naturalmente la
stessa equivalenza si ha anche nel processo di respirazione.
La Fig. 3 mostra il risultato di misure di questo tipo per un abete di
Douglas in un giorno primaverile e in un giorno estivo. Si noti che
ovviamente di notte il flusso di CO![]() è negativo, perchè è
operante solo la respirazione. La misura del flusso di CO
è negativo, perchè è
operante solo la respirazione. La misura del flusso di CO![]() di giorno
è indicativo della produzione primaria netta, in quanto sia la
fotosintesi sia la respirazione sono presenti. Le misure notturne permettono
di valutare l'energia persa per sola respirazione e di risalire quindi alla
produzione primaria lorda. Da notare anche l'influenza dell'intensità
luminosa e della temperatura.
di giorno
è indicativo della produzione primaria netta, in quanto sia la
fotosintesi sia la respirazione sono presenti. Le misure notturne permettono
di valutare l'energia persa per sola respirazione e di risalire quindi alla
produzione primaria lorda. Da notare anche l'influenza dell'intensità
luminosa e della temperatura.
![\includegraphics[scale=1]{abete}](img162.gif) |
Le misure dirette di scambio del carbonio con l'atmosfera sono comunque
piuttosto sofisticate in quanto richiedono l'uso di strumentazione
complicata o di isotopi quali ![]() C. Il metodo più semplice di misura
della PPN è quello che fa ricorso al rilevamento e al bilancio degli
incrementi e delle perdite di biomassa della foresta o della prateria di cui
si voglia misurare la produzione primaria. Più precisamente indichiamo
con [
C. Il metodo più semplice di misura
della PPN è quello che fa ricorso al rilevamento e al bilancio degli
incrementi e delle perdite di biomassa della foresta o della prateria di cui
si voglia misurare la produzione primaria. Più precisamente indichiamo
con [![]() ,
, ![]() ] l'intervallo di tempo relativamente al quale vogliamo
misurare la PPN. Con opportuni metodi si possono rilevare le biomasse dei
produttori primari al tempo
] l'intervallo di tempo relativamente al quale vogliamo
misurare la PPN. Con opportuni metodi si possono rilevare le biomasse dei
produttori primari al tempo ![]() e al tempo
e al tempo ![]() :
: ![]() e
e ![]() . La
produzione primaria netta non è altro che l'energia che viene utilizzata
nel periodo [
. La
produzione primaria netta non è altro che l'energia che viene utilizzata
nel periodo [![]() ,
, ![]() ] per sintetizzare nuova biomassa. Si potrebbe
perciò pensare che l'equivalente in biomassa della PPN è
] per sintetizzare nuova biomassa. Si potrebbe
perciò pensare che l'equivalente in biomassa della PPN è
![]() . Ma questo sarebbe vero solo se in tale periodo non ci
fossero perdite di biomassa dovute alla morte di piante o di parti di
piante. Poiché a causa dell'azione degli erbivori, del prelievo da parte
dell'uomo e dei naturali processi di invecchiamento tali perdite
si verificano, bisogna tenere conto che una parte della PPN va semplicemente
a rimpiazzare la biomassa morta. Perciò per un corretto calcolo della
PPN si deve anche misurare quanta biomassa viene persa per mortalità
durante l'intervallo di tempo considerato:
. Ma questo sarebbe vero solo se in tale periodo non ci
fossero perdite di biomassa dovute alla morte di piante o di parti di
piante. Poiché a causa dell'azione degli erbivori, del prelievo da parte
dell'uomo e dei naturali processi di invecchiamento tali perdite
si verificano, bisogna tenere conto che una parte della PPN va semplicemente
a rimpiazzare la biomassa morta. Perciò per un corretto calcolo della
PPN si deve anche misurare quanta biomassa viene persa per mortalità
durante l'intervallo di tempo considerato:
![]() . In definitiva
la produzione primaria netta in termini di biomassa si ottiene come
. In definitiva
la produzione primaria netta in termini di biomassa si ottiene come
Tale valore può essere convertito in kcal mediante la valutazione a mezzo di bomba calorimetrica del contenuto calorico medio della biomassa dei produttori. Così pure si può convertire tale valore in moli di anidride carbonica, come visto precedentemente, o equivalentemente in grammi di carbonio. Per gran parte dei tessuti delle piante il contenuto di carbonio è circa il 45-50% del peso secco (cioè della biomassa privata di acqua). Perciò i valori di PPN espressi in peso secco sono di solito un po' più del doppio di quelli espressi in carbonio.
Negli ambienti acquatici il metodo di gran lunga più usato per misurare la produzione primaria è quello che rileva le variazioni delle concentrazioni di ossigeno. Infatti, mentre nell'atmosfera l'ossigeno è presente in quantità enormi e i processi di fotosintesi e di respirazione non sono in grado di farne variare apprezzabilmente la concentrazione, non così è nell'acqua dove le concentrazioni di ossigeno disciolto sono estremamente basse e possono venire influenzate in modo rilevante dalla fotosintesi di alghe e piante acquatiche e dalla respirazione degli organismi acquatici, siano essi vegetali o animali. L'idea perciò è di misurare i flussi di ossigeno sapendo che la produzione di 6 moli di ossigeno nel processo fotosintetico equivale all'assorbimento di 709 kcal. In pratica si utilizza il cosiddetto metodo delle bottiglie chiara e scura. Ad una certa profondità vengono presi campioni d'acqua contenenti fitoplankton e viene misurata la concentrazione di ossigeno. Tali campioni vengono poi immediatamente sigillati in bottiglie chiare e scure e risospesi alla medesima profondità (cioè con le medesime condizioni di temperatura e di intensità luminosa). Nelle bottiglie chiare avviene ovviamente sia la fotosintesi sia la respirazione, in quelle scure solo la respirazione. Dopo un certo tempo le bottiglie vengono ricuperate e si misura la variazione della concentrazione di ossigeno avvenuta. Il cambiamento di concentrazione di ossigeno nelle bottiglie chiare fornisce una misura della produzione netta, non del tutto precisa in quanto include la respirazione anche di batteri e zooplankton (eventuali organismi più grossi vengono di solito eliminati!); il cambiamento di concentrazione nella bottiglia scura fornisce una misura della respirazione delle alghe, dei batteri e dello zooplankton. Sommando la produzione netta calcolata dalla bottiglia chiara alla respirazione calcolata dalla bottiglia scura si ottiene la produzione primaria lorda, stimata con buona precisione perchè le due respirazioni, nella bottiglia chiara e in quella scura, anche se non riferite solo ai produttori primari, si elidono reciprocamente.
 |
Con questi metodi di stima della produzione primaria è possibile stabilire come varia la PPN a seconda dei diversi tipi di vegetazione presenti nelle varie parti del nostro pianeta. La Tab. 1 riporta alcuni dati medi a questo proposito affiancandoli ai valori di biomassa. È interessante notare come la PPN in ambiente terrestre raggiunga i suoi valori massimi nella foresta pluviale tropicale e vada decrescendo progressivamente verso i poli. Tuttavia le praterie e le tundre sono meno produttive delle foreste presenti alle medesime latitudini. La produzione primaria dell'oceano aperto è molto bassa, paragonabile a quella della tundra artica e dei deserti, ma le zone di mare prossime alla terraferma, e in particolare estuari e lagune, possono essere molto produttive, più del terreno coltivato. Si noti però che l'oceano aperto ha il più alto rapporto tra PPN e biomassa che la produce: il fitoplankton infatti non ha strutture di sostegno e tutta la biomassa concorre al processo di fotosintesi.
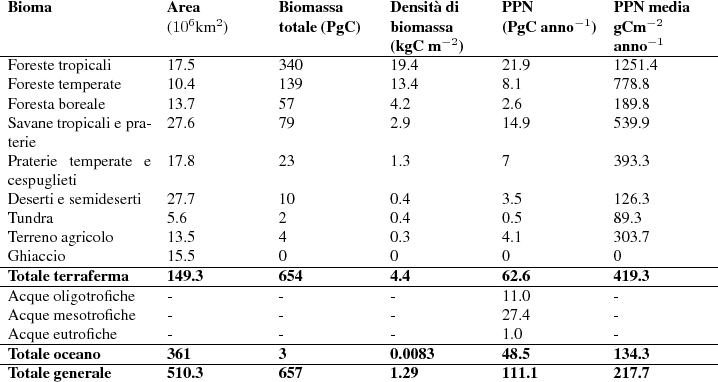 |
È anche interessante stabilire delle stime complessive (riferite cioè
all'intero globo terracqueo) della produzione primaria netta e della
biomassa e stabilire in quale proporzione i vari ecosistemi concorrono al
bilancio globale. Esistono diverse stime a livello del nostro pianeta basate
su compilazioni di dati di diversa fonte. La compilazione storicamente più citata
è quella di Whittaker e Likens (1973), ormai superata perché la stima della biomassa
vegetale sulla terraferma è eccessiva (più di
800![]() 10
10![]() gC). Stime successive (Olson et al., 1983) fornirono
valori più bassi per quanto riguarda la biomassa delle piante terrestri
(che costituiscono comunque la quasi totalità della biomassa vegetale):
560
gC). Stime successive (Olson et al., 1983) fornirono
valori più bassi per quanto riguarda la biomassa delle piante terrestri
(che costituiscono comunque la quasi totalità della biomassa vegetale):
560![]() 10
10![]() gC. Nella Tab. 2 vengono riassunte le informazioni
più recenti. Come si
può notare la PPN globale si aggira sui 110
gC. Nella Tab. 2 vengono riassunte le informazioni
più recenti. Come si
può notare la PPN globale si aggira sui 110![]() 10
10![]() gC anno
gC anno![]() , mentre la biomassa totale vegetale è stimata in più di
650
, mentre la biomassa totale vegetale è stimata in più di
650![]() 10
10![]() gC.
È interessante notare come la produzione primaria
netta per unità di area negli ecosistemi marini è decisamente
inferiore a quella degli ecosistemi terrestri, ma, a causa della maggiore
estensione dei mari rispetto alla terraferma, la PPN totale è di poco
inferiore a quella degli ecosistemi terrestri. Tale PPN, come già fatto
osservare, è poi ottenuta con una biomassa che è circa lo 0.46% di
quella delle piante terrestri.
gC.
È interessante notare come la produzione primaria
netta per unità di area negli ecosistemi marini è decisamente
inferiore a quella degli ecosistemi terrestri, ma, a causa della maggiore
estensione dei mari rispetto alla terraferma, la PPN totale è di poco
inferiore a quella degli ecosistemi terrestri. Tale PPN, come già fatto
osservare, è poi ottenuta con una biomassa che è circa lo 0.46% di
quella delle piante terrestri.
Ci si può chiedere qual è l'efficienza delle piante nel catturare
l'energia solare mediante il processo di fotosintesi clorofilliana.
Estremamente bassa, come permette di appurare un calcolo approssimativo.
Ogni anno la radiazione solare nelle lunghezze d'onda corte, cioè quelle
sfruttabili dalle piante, che raggiunge la superficie terrestre, inclusi gli
oceani, è di circa 3![]() 10
10![]() J (Harte, 1985). Dalla Tab. 2
risulta che la PPN totale annuale è di 111.1
J (Harte, 1985). Dalla Tab. 2
risulta che la PPN totale annuale è di 111.1![]() 10
10![]() gC. Abbiamo
visto che lo scambio di 72 gC corrisponde a 709 kcal = 4.18
gC. Abbiamo
visto che lo scambio di 72 gC corrisponde a 709 kcal = 4.18![]() 0.709
0.709![]() 10
10![]() J (ricordatevi che 1 cal equivale a 4.18 J!) e
perciò con facili calcoli si ottiene che la PPN totale annuale è di
4.57
J (ricordatevi che 1 cal equivale a 4.18 J!) e
perciò con facili calcoli si ottiene che la PPN totale annuale è di
4.57![]() 10
10![]() J. Ne viene quindi che solo lo 0.15%della
radiazione incidente utile è accumulato nella biomassa delle piante!
Naturalmente una certa parte della radiazione solare viene a cadere su zone
prive di vegetazione, ma nonostante questo la percentuale di energia solare
catturata dalle piante rimane comunque molto bassa. Per la precisione
possiamo definire due tipi di efficienze:
J. Ne viene quindi che solo lo 0.15%della
radiazione incidente utile è accumulato nella biomassa delle piante!
Naturalmente una certa parte della radiazione solare viene a cadere su zone
prive di vegetazione, ma nonostante questo la percentuale di energia solare
catturata dalle piante rimane comunque molto bassa. Per la precisione
possiamo definire due tipi di efficienze:
Efficienza fotosintetica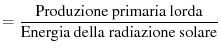 |
|||
Efficienza di produzione netta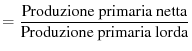 |
![\includegraphics[scale=1]{product}](img176.gif) |
Che cosa determina i diversi valori di produzione primaria che caratterizzano i diversi ecosistemi, come messo in evidenza dalle Tab. 1 e 2? Nonostante che l'energia immagazzinata nelle piante sia in definitiva quella fornita dal sole, non è in generale la luce ad essere il fattore limitante che controlla la produzione primaria. Se così fosse, i mari tropicali dovrebbero essere massimamente produttivi e quelli polari estremamente improduttivi, come mostra la Fig. 4. Invece il Mar dei Sargassi ha una PPN molto bassa, mentre l'Oceano Antartico è la regione oceanica più produttiva. Il fatto è che per produrre biomassa vegetale non basta energia. In primo luogo il processo di fotosintesi clorofilliana ha bisogno di acqua, che è decisamente deficiente in alcuni climi caratterizzati da grande intensità della radiazione solare. Ma in più ci vogliono tutta una serie di elementi o composti che non possono venire acquisiti con la fotosintesi (la quale, ricordiamoci, fornisce alla pianta C, H, O) e che sono mattoni fondamentali della materia vivente. I principali elementi che le piante devono procurarsi assorbendoli dal suolo, dall'atmosfera o, se acquatiche, dall'acqua sono azoto, fosforo e zolfo. Altri elementi come silicio o ferro o manganese sono importanti, ma di solito non costituiscono quantitativamente una parte rilevante della biomassa e sono presenti nell'ambiente in quantità sufficienti ai bisogni dei produttori primari.