Gli ecosistemi insulari sono sempre stato oggetto di particolare attenzione da parte di naturalisti ed ecologi. Infatti la ovvia discontinuità terra-acqua pone dei limiti ben precisi alla distribuzione delle specie rendendo le comunità insulari sostanzialmente chiuse ad interazioni ecologiche con l'esterno. La diversità nelle isole ha delle caratteristiche molto interessanti.
![\includegraphics[width=0.75\linewidth]{galapagos}](img623.gif) |
La prima di queste caratteristiche può essere illustrata con riferimento al caso dell'arcipelago delle Galapagos il quale, a partire dalla famosa visita di Darwin nel 1835, è sempre stato oggetto di interesse da parte dei naturalisti per la sua speciale fauna. La Fig. 6 riporta, in funzione dell'area delle diverse isole, il numero delle piante terrestri che si trovano in ciascuna isola. Si può notare che in scala bilogaritmica tale relazione è approssimativamente lineare. In altri termini si ha
Un fatto interessante notato da Preston (1962) è che l'esponente ![]() della
relazione numero di specie-area dell'isola tende ad essere circa 0.3 per
moltissime situazioni, dalle formiche della Melanesia ai vertebrati delle
isole del Lago Michigan agli scarabei delle Indie Occidentali.
della
relazione numero di specie-area dell'isola tende ad essere circa 0.3 per
moltissime situazioni, dalle formiche della Melanesia ai vertebrati delle
isole del Lago Michigan agli scarabei delle Indie Occidentali.
![\includegraphics[scale=1]{rettili}](img627.gif) |
La seconda caratteristica della diversità delle isole ci può anche
illuminare sulle probabili cause di ciò che osserviamo: è il
cosiddetto effetto distanza, che può essere facilmente illustrato dall'esempio di
Fig. 8, basato sui dati raccolti da Diamond (1973)
sugli uccelli dell'arcipelago della Nuova Guinea. In tale figura
vengono riportati solo i dati relativi alle isole che distano più di 500
km dalla terraferma (cioè dall'isola di Nuova Guinea propriamente
detta). Sull'asse delle ascisse abbiamo la distanza dalla Nuova Guinea (in
scala lineare); sull'asse delle ordinate abbiamo (in scala logaritmica) il
rapporto tra il numero effettivo di uccelli che si trovano su una certa
isola e il numero di uccelli che ci si aspetterebbe di avere su un'isola
della stessa area, ma distante dalla terraferma meno di 500 km. In altri
termini, Diamond tarò la relazione ![]() sulle isole vicine alla
nuova Guinea e poi mise in evidenza che, per le isole lontane più di 500
km, si ha un effetto distanza in quanto il rapporto diminuisce con la
distanza dalla terraferma. In sintesi, possiamo perciò dire che il numero di specie
su un'isola è funzione crescente dell'area e funzione decrescente della
distanza dalla terraferma.
sulle isole vicine alla
nuova Guinea e poi mise in evidenza che, per le isole lontane più di 500
km, si ha un effetto distanza in quanto il rapporto diminuisce con la
distanza dalla terraferma. In sintesi, possiamo perciò dire che il numero di specie
su un'isola è funzione crescente dell'area e funzione decrescente della
distanza dalla terraferma.
![\includegraphics[width=0.5\linewidth]{effettivo}](img629.gif) |
L'effetto distanza suggerisce che sia l'immigrazione dalla terraferma (tanto più difficile quanto maggiore è la distanza) a rifornire di specie le isole. Ed è proprio sul bilancio tra immigrazione di nuove specie ed estinzione di quelle già presenti che è basata la teoria più nota per spiegare la diversità delle comunità insulari: quella dovuta a MacArthur e Wilson (1967) ed esposta in dettaglio nel famoso libro ``The theory of island biogeography''. Secondo questi autori il numero di specie presente su di un'isola varia come risultato di due forze contrapposte: da una parte specie non ancora presenti sull'isola possono giungere sull'isola dalla terraferma (non solo gli uccelli e gli insetti o le piante dotate di semi anemofili, ma anche altri organismi possono immigrare su un'isola portati da "zattere" naturali o artificiali), dall'altra le specie già presenti possono estinguersi. Come mai si ha questa possibilità di estinzione? Semplicemente perchè le popolazioni delle specie insulari sono in generale molto più piccole di quelle ospitate dalla terraferma e le popolazioni di piccole dimensioni sono soggette a una serie di problemi:
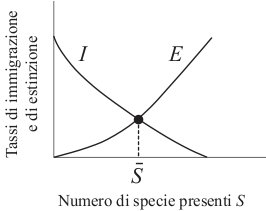 |
La situazione è perciò quella descritta nella
Fig. 9. Esiste un valore in corrispondenza del
quale si uguagliano il tasso di immigrazione e il tasso di estinzione. Se
![]() è inferiore a
è inferiore a ![]() , il numero di specie sull'isola tende a crescere,
perché
, il numero di specie sull'isola tende a crescere,
perché ![]() , se
, se ![]() è superiore a
è superiore a ![]() , il numero di specie sull'isola tende
a diminuire, perché
, il numero di specie sull'isola tende
a diminuire, perché ![]() . Perciò
. Perciò ![]() è una condizione di equilibrio
stabile che dovrebbe caratterizzare tutte quelle isole in cui non ci sono
stati recentemente importanti disturbi ambientali. Secondo MacArthur e
Wilson il numero di specie che si contano su di un'isola dovrebbe essere
dunque approssimativamente uguale a
è una condizione di equilibrio
stabile che dovrebbe caratterizzare tutte quelle isole in cui non ci sono
stati recentemente importanti disturbi ambientali. Secondo MacArthur e
Wilson il numero di specie che si contano su di un'isola dovrebbe essere
dunque approssimativamente uguale a ![]() . Va notato che
. Va notato che ![]() è un equilibrio per
così dire dinamico, perché quello che rimane costante è il numero
di specie, non l'identità delle specie: infatti, in corrispondenza di
è un equilibrio per
così dire dinamico, perché quello che rimane costante è il numero
di specie, non l'identità delle specie: infatti, in corrispondenza di ![]() ,
sia il tasso di immigrazione sia quello di estinzione sono positivi (e
uguali), non nulli. C'è quindi un ricambio delle specie presenti
sull'isola, che nell'unità di tempo è pari proprio a
,
sia il tasso di immigrazione sia quello di estinzione sono positivi (e
uguali), non nulli. C'è quindi un ricambio delle specie presenti
sull'isola, che nell'unità di tempo è pari proprio a
![]() .
.
Una conferma della teoria di MacArthur e Wilson venne da alcuni interessanti esperimenti condotti da Simberloff e dal medesimo Wilson in quattro piccolissime isole a mangrovia (il diametro varia tra gli 11 e i 18 metri) vicino alla Florida (Simberloff e Wilson, 1970). Queste isolette erano popolate esclusivamente da una fauna di artropodi (insetti, ragni, scorpioni, ecc.). Il numero di specie sulla terraferma è di circa un migliaio, mentre ogni isoletta ospitava dalle venti alle quaranta specie. Simberloff e Wilson sterminarono per mezzo di bromuro di metile l'intera fauna delle quattro isolette e poi osservarono il processo di ricolonizzazione delle quattro isole. I risultati sono riportati in Fig. 10 e mettono in evidenza i seguenti fatti:
![\includegraphics[width=0.75\linewidth]{simberloff}](img641.gif) |
Vediamo ora come mediante la teoria di MacArthur e Wilson si possono
giustificare l'effetto area e l'effetto distanza. Consideriamo dapprima due
isole alla stessa distanza dalla terraferma, di cui una piccola,
caratterizzata da un tasso di immigrazione ![]() e di estinzione
e di estinzione
![]() , e l'altra grande, caratterizzata da
, e l'altra grande, caratterizzata da ![]() e
e ![]() . A
parità di specie presenti l'isola grande sarà caratterizzata da un
minore tasso di estinzione, perchè offrirà più risorse alle
popolazioni che ospita, le quali saranno perciò più abbondanti e con
minore sovrapposizione delle nicchie ecologiche. Per quanto riguarda il
tasso di immigrazione, esso non è sostanzialmente influenzato dalla
grandezza dell'isola, anche se possiamo pensare che un'isola più grossa
sia più facilmente rinvenibile da un organismo migrante che un'isola
piccola. In sintesi abbiamo
. A
parità di specie presenti l'isola grande sarà caratterizzata da un
minore tasso di estinzione, perchè offrirà più risorse alle
popolazioni che ospita, le quali saranno perciò più abbondanti e con
minore sovrapposizione delle nicchie ecologiche. Per quanto riguarda il
tasso di immigrazione, esso non è sostanzialmente influenzato dalla
grandezza dell'isola, anche se possiamo pensare che un'isola più grossa
sia più facilmente rinvenibile da un organismo migrante che un'isola
piccola. In sintesi abbiamo
Queste condizioni sono sufficienti, come facilmente dimostrato dalla Fig. 11a, a dedurre che all'equilibrio l'isola più piccola deve ospitare meno specie.
Consideriamo adesso due isole della medesima area, ma a diversa distanza
dalla terraferma. L'isola più vicina è caratterizzata da tassi
![]() e
e ![]() , l'isola più lontana da tassi
, l'isola più lontana da tassi ![]() e
e
![]() . Non c'è alcuna ragione di pensare che i tassi di estinzione
sulle due isole siano diversi, mentre chiaramente il tasso di immigrazione
nell'isola più lontana è minore:
. Non c'è alcuna ragione di pensare che i tassi di estinzione
sulle due isole siano diversi, mentre chiaramente il tasso di immigrazione
nell'isola più lontana è minore:
La Fig. 11b mostra chiaramente anche per questo caso che all'equilibrio l'isola più lontana deve albergare meno specie.
![\includegraphics[scale=1]{vicini_lontani}](img654.gif) |