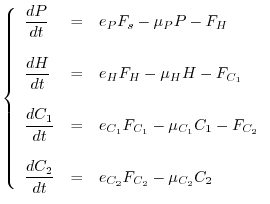Nel Capitolo 4 si è già messo in luce che l'energia entrante in un ecosistema come luce solare, catturata dalla vegetazione col processo di fotosintesi, va rapidamente dissipandosi lungo le catene trofiche, poiché le efficienze ecologiche sono piuttosto piccole. Questo fenomeno fondamentale ha conseguenze notevoli sulla struttura delle comunità. Già Elton (1927) aveva fatto notare che, se ciascun livello trofico viene rappresentato con un blocco di dimensione pari alla produttività del livello (cioè alla sua produzione netta per unità di area), si ottiene una tipica struttura piramidale con alla base i produttori primari e in cima i carnivori dominanti. Infatti, poiché la produzione di un livello va parzialmente a rimpiazzare anche la biomassa morta per predazione da parte del livello più alto e poiché l'efficienza di produzione lorda non supera quasi mai il 30% ed è spesso inferiore a tale valore, ne consegue che il rapporto tra le produzioni nette di due livelli successivi varia di solito tra il 5 e il 20%.
È invalso l'uso di rappresentare in maniera graficamente analoga anche i numeri di individui e le biomasse corrispondenti a ciascun livello trofico. Generalmente si ottengono ancora delle piramidi (piramidi ecologiche) come è mostrato in Fig. 5. Va tuttavia notato che l'utilizzo dei numeri di individui riferiti ad un certo livello trofico non ha alcun senso dal punto di vista energetico: se i produttori primari sono piante ad alto fusto saranno poche di numero, ma con grande biomassa individuale ed elevata produttività, se sono alghe unicellulari saranno in numero sterminato, ma con piccola biomassa individuale, e lo stesso inconveniente si verifica anche per gli altri livelli trofici. L'esempio della foresta temperata in Fig. 5 chiarisce questo semplice fatto. Più sensato è utilizzare le piramidi di biomassa, poichè l'accumulo di biomassa corrisponde certamente ad un accumulo di energia.
![\includegraphics[width=1\linewidth]{piramidi}](img451.gif) |
Va però messo in evidenza che non necessariamente la quantità di energia accumulata nei vari livelli va diminuendo passando dai produttori primari ai carnivori. Un'analogia idraulica chiarisce perchè: un lago molto grande può avere un immissario e un emissario con piccole portate e d'altra parte un lago più piccolo può avere un immissario ed un emissario con grandi portate (pensate come esempio al Lago di Garda e al Lago Maggiore!). Perciò non necessariamente un livello trofico basso, caratterizzato da elevati flussi di energia entranti e uscenti è anche caratterizzato da elevata biomassa. Tuttavia, come per i laghi esiste comunque una certa correlazione tra il volume del loro invaso e il flusso degli immissari ed emissari, così per le comunità ecologiche esiste una certa correlazione tra le biomasse dei diversi livelli trofici e le loro produttività. Perciò, come mostra la Fig. 5, raramente le piramidi di biomassa sono rovesciate ovvero si ha meno biomassa ai livelli trofici inferiori che non a quelli superiori. Una tipica eccezione è quella del plankton (vedi in Fig. 5 il caso del Lago Maggiore in inverno) in cui la biomassa del fitoplankton può essere inferiore a quella dello zooplankton, perché esso viene consumato molto rapidamente dallo zooplankton erbivoro. L'esempio di Silver Springs mostra una sintonia tra la piramide di biomassa e quella di produttività, ma il rapporto tra flusso energetico entrante in un comparto e dimensione del comparto è comunque molto variabile.
Gli esempi di Fig. 5 confermano anche quanto affermato nel precedente paragrafo, ovvero una certa disattenzione verso il sistema di detrito che invece è presente in tutte le comunità e svolge un ruolo fondamentale nel riciclo dei nutrienti. Solo in pochi casi viene riportata la dimensione del comparto dei decompositori e il flusso di energia in esso entrante. Più in generale invece la struttura trofica completa di una comunità è, perlomeno approssimativamente, quella mostrata in Fig. 6. In essa vengono indicati con rettangoli i comparti di accumulo dell'energia e con frecce i flussi di energia tra i comparti. In omaggio alla tradizione il sistema di produttività di piante, erbivori, carnivori di primo e secondo livello (costituito dalle catene di pascolo) viene graficamente mostrato come una piramide e così pure il sistema dei decompositori e detritivori (costituito dalle catene di detrito).
![\includegraphics[scale=0.8]{struttura}](img452.gif) |
Si noti che le vie lungo cui scorre l'energia sono sostanzialmente le stesse nel sistema di pascolo e nel sistema di detrito con un'eccezione fondamentale: la biomassa morta è definitivamente persa per la catena di pascolo ed entra nel sistema dei decompositori. Poiché ad ogni passaggio da un livello trofico a quello superiore una buona parte dell'energia va persa sotto forma di calore nel processo di respirazione, ne consegue che l'energia che entra nel sistema di detrito come biomassa morta viene alla fine completamente metabolizzata e persa attraverso la respirazione come calore, anche se questo può richiedere alcuni ricicli nel sistema dei decompositori. Nel sistema di pascolo non ci sono invece ricicli dell'energia e questa è una delle ragioni per cui esso è più facilmente studiato di quello di detrito.
Sulla base dello schema di Fig. 6 è possibile impostare un bilancio energetico della comunità. Per semplicità limitiamo la nostra attenzione al sistema di pascolo. Si tratta di scrivere per ogni comparto il seguente bilancio nell'unità di tempo:
| Energia immagazzinata | = | Flusso di energia entrante - Flusso di energia uscente |
| = | tempo | |
| = | contenuti energetici dei rispettivi comparti | |
| = | flusso di energia proveniente dal sole | |
| = | efficienza totale della produzione primaria (PPN/ |
|
| = | flusso di energia acquisito dal comparto |
|
| = | efficienza di produzione lorda del comparto |
|
| = | tasso di mortalità del comparto |
|
| . |
Spesso è ragionevole supporre che la comunità si trovi all'equilibrio e che valgano quindi le seguenti relazioni tra i vari flussi energetici
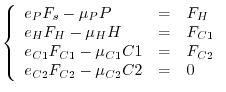
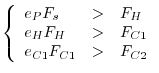
![[*]](crossref.gif) abbiamo indicato come ingestione) vanno diminuendo rapidamente lungo la catena di pascolo
(struttura piramidale dei flussi di energia). Per la medesima ragione vanno
diminuendo le produzioni nette
abbiamo indicato come ingestione) vanno diminuendo rapidamente lungo la catena di pascolo
(struttura piramidale dei flussi di energia). Per la medesima ragione vanno
diminuendo le produzioni nette
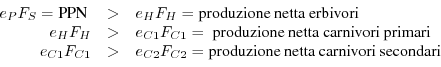
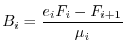
Naturalmente i bilanci energetici possono essere fatti anche ad un livello più dettagliato, scindendo i comparti in sottocomparti. La Fig. 7 riporta in forma grafica il bilancio energetico per uno stagno salmastro della Georgia (USA).
![\includegraphics[width=1\linewidth]{georgia}](img476.gif) |
Nonostante che i bilanci energetici varino da caso a caso e che gli studi si siano prevalentemente rivolti al sistema di pascolo delle comunità analizzate, è tuttavia possibile fare qualche generalizzazione riferendosi alle caratteristiche più grossolane di alcuni tipici ecosistemi. La Fig. 8 riassume in maniera grafica le principali tra queste caratteristiche. Vengono mostrate le strutture trofiche di una ipotetica foresta, di una prateria, di una comunità planctonica marina o di un lago di grandi dimensioni e di una comunità di acqua corrente o di un piccolo lago. Per semplicità i comparti sono ridotti a quattro: i produttori primari con i relativi consumatori del sistema di pascolo e la materia organica morta con i relativi consumatori del sistema di detrito. Le grandezze dei rettangoli indicano l'entità dell'accumulo nei comparti, mentre le dimensioni delle frecce indicano l'entità dei flussi energetici tra i vari comparti o con l'ambiente esterno. Le dimensioni di rettangoli e frecce non sono assolute, ma valgono all'interno della singola comunità. Ad esempio i rettangoli dei produttori primari nella foresta e nella prateria hanno più o meno la stessa dimensione non perché abbiano gli stessi valori assoluti (sono ben diversi come sappiamo dalla tabella riportata nei precedenti capitoli), ma per ragioni grafiche. Il fatto importante è la diversa importanza relativa dei comparti e dei flussi. Il comparto della materia organica morta è molto più consistente di quello dei produttori primari nella prateria rispetto alla foresta e nella comunità d'acqua corrente rispetto alla comunità planctonica marina. Nella prateria rispetto alla foresta gli eterotrofi del sistema di pascolo da una parte consumano una maggiore percentuale della produzione primaria netta e dall'altra provvedono un più consistente flusso di materia organica morta. I consumatori del sistema di pascolo hanno la loro massima importanza nelle comunità planctoniche sia in termini di biomassa del comparto rispetto agli altri comparti sia in termini di flussi respiratori. Al contrario il sistema di pascolo è praticamente irrilevante nelle comunità d'acqua corrente dove la produzione primaria netta è assai bassa. Si noti che per le comunità d'acqua corrente è stato messo in evidenza un fattore finora trascurato, ma che non può esserlo per fiumi o piccoli laghi: l'apporto di materiale organico dall'ambiente terrestre, in particolare dal bacino cui il corpo d'acqua appartiene. In questo caso il flusso fondamentale di energia entrante nell'ecosistema non è fornito dagli autotrofi attraverso il processo di fotosintesi, ma è costituito da tale apporto esterno. Nella realtà importazioni ed esportazioni di materia (e quindi di energia) sono presenti in tutti gli ecosistemi. La loro importanza dipende molto da come sono stati definiti i confini dell'ecosistema sotto studio, come abbiamo già discusso nel capitolo 3.
![\includegraphics[width=0.75\linewidth]{trofiche}](img477.gif) |